The structural biochemistry of Zucchini implicates it as a nuclease in piRNA biogenesis
Abstract
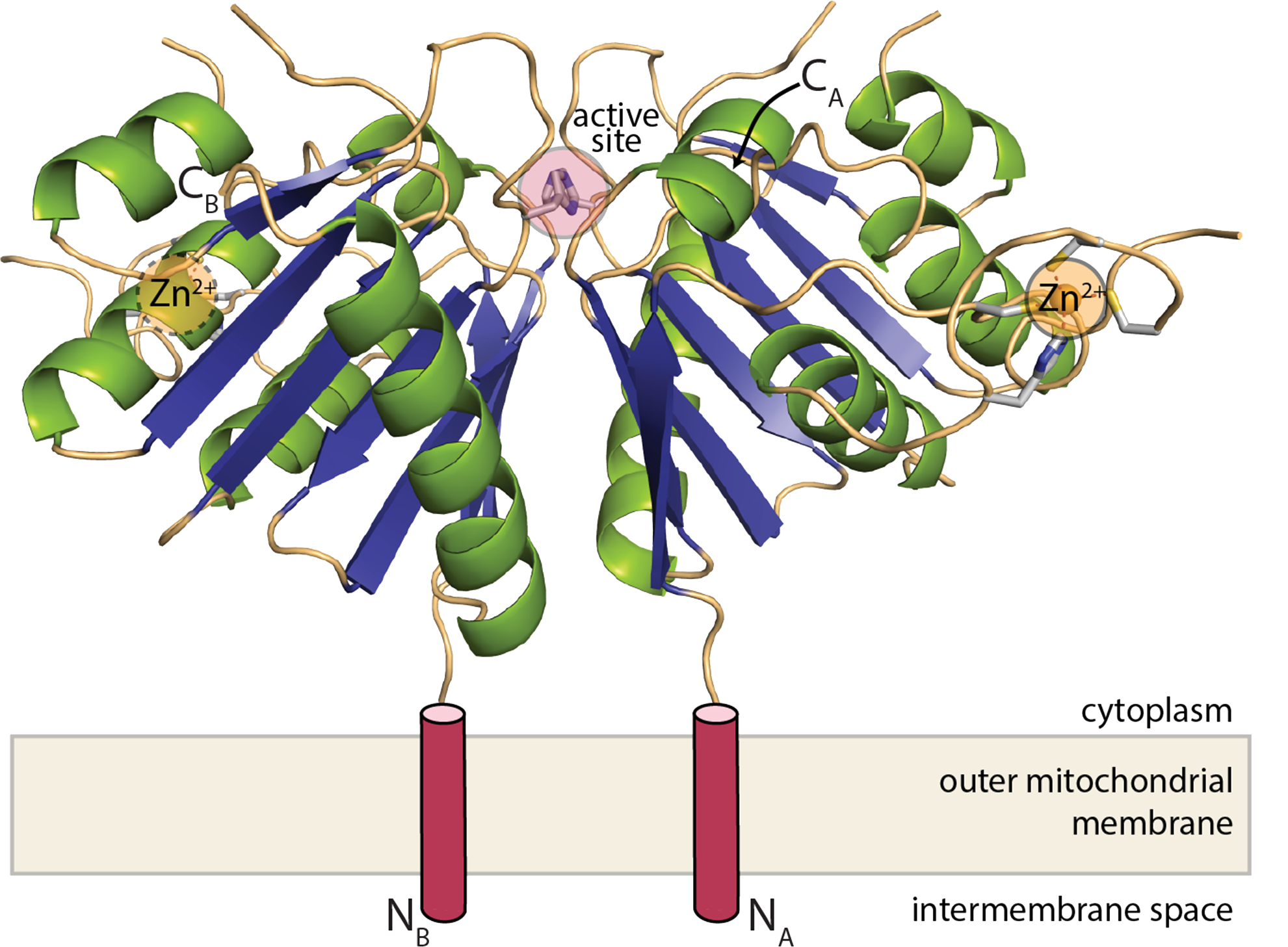 PIWI-family proteins and their associated small RNAs (piRNAs) act in an evolutionarily conserved innate immune mechanism to provide essential protection for germ-cell genomes against the activity of mobile genetic elements. piRNA populations comprise a molecular definition of transposons, which permits them to distinguish transposons from host genes and selectively silence them. piRNAs can be generated in two distinct ways, forming either primary or secondary piRNAs. Primary piRNAs come from discrete genomic loci, termed piRNA clusters, and seem to be derived from long, single-stranded precursors. The biogenesis of primary piRNAs involves at least two nucleolytic steps. An unknown enzyme cleaves piRNA cluster transcripts to generate monophosphorylated piRNA 5’ ends. piRNA 3’ ends are probably formed by exonucleolytic trimming, after a piRNA precursor is loaded into its PIWI partner. Secondary piRNAs arise during the adaptive ‘ping-pong’ cycle, with their 5’ termini being formed by the activity of PIWIs themselves. A number of proteins have been implicated genetically in primary piRNA biogenesis. One of these, Drosophila melanogaster Zucchini, is a member of the phospholipase-D family of phosphodiesterases, which includes both phospholipases and nucleases. Here we produced a dimeric, soluble fragment of the mouse Zucchini homologue (mZuc; also known as PLD6) and show that it possesses single-strand-specific nuclease activity. A crystal structure of mZuc at 1.75 Å resolution indicates greater architectural similarity to phospholipase-D family nucleases than to phospholipases. Together, our data suggest that the Zucchini proteins act in primary piRNA biogenesis as nucleases, perhaps generating the 5’ ends of primary piRNAs.
PIWI-family proteins and their associated small RNAs (piRNAs) act in an evolutionarily conserved innate immune mechanism to provide essential protection for germ-cell genomes against the activity of mobile genetic elements. piRNA populations comprise a molecular definition of transposons, which permits them to distinguish transposons from host genes and selectively silence them. piRNAs can be generated in two distinct ways, forming either primary or secondary piRNAs. Primary piRNAs come from discrete genomic loci, termed piRNA clusters, and seem to be derived from long, single-stranded precursors. The biogenesis of primary piRNAs involves at least two nucleolytic steps. An unknown enzyme cleaves piRNA cluster transcripts to generate monophosphorylated piRNA 5’ ends. piRNA 3’ ends are probably formed by exonucleolytic trimming, after a piRNA precursor is loaded into its PIWI partner. Secondary piRNAs arise during the adaptive ‘ping-pong’ cycle, with their 5’ termini being formed by the activity of PIWIs themselves. A number of proteins have been implicated genetically in primary piRNA biogenesis. One of these, Drosophila melanogaster Zucchini, is a member of the phospholipase-D family of phosphodiesterases, which includes both phospholipases and nucleases. Here we produced a dimeric, soluble fragment of the mouse Zucchini homologue (mZuc; also known as PLD6) and show that it possesses single-strand-specific nuclease activity. A crystal structure of mZuc at 1.75 Å resolution indicates greater architectural similarity to phospholipase-D family nucleases than to phospholipases. Together, our data suggest that the Zucchini proteins act in primary piRNA biogenesis as nucleases, perhaps generating the 5’ ends of primary piRNAs.